|
—Ә—р
1992”N “ҢӢһҗ…ҺY‘еҠwҗ…ҺYҠw•”‘ІӢЖ
1997”N җVҠғ‘еҠw‘еҠwү@Һ©‘RүИҠwҢӨӢҶүИ”ҺҺmҢгҠъүЫ’цҸC—№Ғ@”ҺҺmҒi—қҠwҒj
1998”N “ъ–{ҠwҸpҗUӢ»үп–ў—ҲҠJ‘сҠwҸpҢӨӢҶҗ„җiҺ–ӢЖҢӨӢҶҲх
1999”N “ъ–{ҠwҸpҗUӢ»үп“Б•КҢӨӢҶҲх
2002”N үИҠwӢZҸpҗUӢ»Һ–ӢЖ’cӮіӮ«ӮӘӮҜҢӨӢҶ21Ғu”FҺҜӮЖҢ`җ¬Ғv—МҲжҗк”CҢӨӢҶҲх
2007”N үF“sӢ{‘еҠwҲв“`ҺqҺАҢұҺ{җЭҸyӢіҺцҒ@Ң»ҚЭӮЙҺҠӮйҒB
ҒңӮНӮ¶ӮЯӮЙ
Ғ@ҡM“ы—ЮӮМҗ«ӮНҗ«җхҗF‘МӮМ‘gӮЭҚҮӮнӮ№ӮЙӮжӮБӮДҢҲӮЯӮзӮкӮДӮўӮйҒBӮ·ӮИӮнӮҝҒAXҗхҗF‘МӮрӮаӮВҗёҺqӮӘҺцҗёӮ·ӮйӮ©ҒAYҗхҗF‘МӮрӮаӮВҗёҺqӮӘҺцҗёӮ·ӮйӮ©ӮЙӮжӮБӮДҒAӮ»ӮМҢВ‘МӮМҗ«ӮНҢҲӮЬӮйӮМӮЕӮ ӮйҒBҸ«—ҲҒAҗё‘ғӮаӮөӮӯӮН—‘‘ғӮЙ•Әү»Ӯ·Ӯйҗ¶җB‘ғӮНҒA”ӯҗ¶ҸүҠъӮН–ў•Әү»ӮЕ‘o•ыӮЙ•Әү»Ӯ·Ӯй”\—НӮрӮаӮБӮДӮўӮйҒBҺцҗёӮМҸuҠФӮЙҢҲӮЬӮБӮҪҗ«ӮЙҸ]ӮБӮДҒA–ў•Әү»җ¶җB‘ғӮНҗё‘ғҒAӮаӮөӮӯӮН—‘‘ғӮЦӮЖ•Әү»ӮөҒAҗ¶җB‘ғӮМҗ«ӮЙҸ]ӮБӮДҢВ‘МӮМҗ«ӮНҢҲ’иӮіӮкӮйҒBҗТ’Е“®•ЁӮМҗ«ӮНҒAҡM“ы—ЮӮМӮжӮӨӮЙҲв“`“IӮЙҢҲӮЯӮзӮкӮйҺнӮӘ‘Ҫҗ”ӮЕӮ ӮйӮЖҺvӮнӮкӮйӮӘҒAаҰ’Һ—ЮӮвӢӣ—ЮӮМҲк•”ӮЙӮНҠВӢ«—vҲцӮЙӮжӮиҢҲӮЯӮзӮкӮйҺнӮаӮ ӮйҒBӮЬӮҪҒAӢӣ—ЮӮЙӮНҲкҗ¶ӮМҠФӮЙҗё‘ғӮрӮаӮВ—YӮМҺһҠъӮЖ—‘‘ғӮрӮаӮВҺ“ӮМҺһҠъӮЖӮрҚsӮ«—ҲӮ·ӮйҺнӮвҒAҗё‘ғӮЖ—‘‘ғӮЖӮр“ҜҺһӮЙӮаӮВҺ“—Y“Ҝ‘МӮМҺнӮа•сҚҗӮіӮкӮДӮўӮйҒBӮұӮМӮжӮӨӮЙӢӣ—ЮӮМҗ«ҢҲ’иҒEҗ«•Әү»ӮМ—lҺqӮНҡM“ы—ЮӮМӮ»ӮкӮЖ”дӮЧӮйӮЖ”сҸнӮЙ‘Ҫ—lӮЕӮ ӮйҒBӮЬӮҪҒAҸгӢLӮМӮжӮӨӮЙҺ©‘RӮЙӢNӮұӮйҗ«“]Ҡ·ӮЙүБӮҰҒAҲв“`“IӮЙҗ«ӮМҢҲӮЯӮзӮкӮДӮўӮй‘ҪӮӯӮМҺнӮЕӮаҗ«ғXғeғҚғCғhғzғӢғӮғ“ҸҲ—қӮЙӮжӮиҒAҗ«ӮрҗlҲЧ“IӮЙ“]Ҡ·ӮіӮ№ӮйӮұӮЖӮӘүВ”\ӮЕӮ ӮйҒB’КҸнҒAӮұӮкӮзҗ«“]Ҡ·ҢВ‘МӮНҗіҸнӮИҗёҺqӮв—‘ӮрӮВӮӯӮйҒBӮұӮМӮжӮӨӮЙҡM“ы—ЮӮЖ”дӮЧӮДҗ«ӮМүВ‘Yҗ«ӮӘ‘еӮ«ӮўӮұӮЖӮӘҒAӢӣ—ЮӮрҗ«ҢҲ’иҒEҗ«•Әү»ӮМғӮғfғӢ“®•ЁӮЖӮөӮДҺg—pӮ·Ӯй—ҳ“_ӮМҲкӮВӮЕӮ ӮйҒB
Ғ@Ҳв“`“IӮЙҗ«ӮӘҢҲ’иӮіӮкӮйҺнӮЙӮНҗ«ҢҲ’иҲв“`ҺqӮӘ‘¶ҚЭӮ·ӮйӮНӮёӮЕӮ ӮйҒBҡM“ы—ЮӮМҗ«ҢҲ’иҲв“`ҺqҒiSRY/SryҒjӮМ“Ҝ’и(1990”N)ҲИ—ҲҒA‘ҪӮӯӮМҢӨӢҶҺТӮӘ‘јӮМҗТ’Е“®•ЁҺнӮЙӮЁӮўӮДҗ«ҢҲ’иҲв“`ҺqӮМ“Ҝ’иӮрҺҺӮЭӮҪҒBӮөӮ©ӮөӮИӮӘӮзҒAҢ»ҚЭӮМӮЖӮұӮлҒAҗ«ҢҲ’иҲв“`ҺqӮЖӮөӮД“Ҝ’иӮіӮкӮДӮўӮйӮМӮНҒA‘е‘Ҫҗ”ӮМ—L‘Щ”ХҡM“ы—ЮӮЙӢӨ’КӮИSRY/SryҲв“`ҺqӮЖҚЕӢЯүдҒXӮӘ“Ҝ’иӮөӮҪғҒғ_ғJӮМDMYҲв“`Һq1)ӮМӮЭӮЕӮ ӮйҒBҡM“ы—ЮӮМҸкҚҮҒAӮwҗхҗF‘МӮЖӮxҗхҗF‘МӮЖӮНҢ`‘ФӮӘ‘еӮ«ӮӯҲЩӮИӮБӮДӮўӮйҒBӮЬӮҪҒAӮxҗхҗF‘М“БҲЩ“IӮИҲв“`ҺqӮӘ‘Ҫҗ”‘¶ҚЭӮ·ӮйҒBӮВӮЬӮиҒAҡM“ы—ЮӮМӮwҗхҗF‘МӮЖӮxҗхҗF‘МӮЖӮНҢ`‘Ф“IӮЙӮаӢ@”\“IӮЙӮа‘еӮ«ӮӯҲЩӮИӮБӮДӮўӮйҒBҲк•ыҒAӢӣ—ЮӮЕӮНҗ«җхҗF‘МӮМҢ`‘ФӮӘҲЩӮИӮйҺнӮаӮ ӮйӮӘҒA‘е‘Ҫҗ”ӮМҺнӮЙӮЁӮўӮДҒAҗ«җхҗF‘МӮНҢ`‘Ф“IӮЙ•Әү»ӮөӮДӮўӮИӮўҒBғҒғ_ғJӮМҸкҚҮҒAҗ«ҢҲ’и—lҺ®ӮНXX-XYҢ^ӮМ—YӮЦғeғҚӮЕӮ ӮйӮӘҒAҗ«“]Ҡ·ҸҲ—қӮЙӮжӮи“ҫӮҪXX—YӮНҗіҸнӮИҗёҺqӮрҒAXYӮвYYӮМҺ“ӮаҗіҸнӮИ—‘ӮрҺYӮЮҒBӮВӮЬӮиҒAҡM“ы—ЮӮМҸкҚҮӮЖҲЩӮИӮиҒAXXҢВ‘МӮаҗё‘ғҢ`җ¬ҒAҗёҺqҢ`җ¬ӮЙ•K—vӮИҲв“`ҺqӮрӮ·ӮЧӮДӮаӮБӮДӮўӮйӮөҒAYYҢВ‘МӮа—‘‘ғҢ`җ¬ҒA—‘Ң`җ¬ӮЙ•K—vӮИҲв“`ҺqӮр‘SӮДҺқӮБӮДӮўӮйӮМӮЕӮ ӮйҒBӮұӮМӮұӮЖӮ©ӮзҒAғҒғ_ғJӮрҠЬӮЯӮД‘ҪӮӯӮМӢӣ—ЮӮЕӮНҒAXҗхҗF‘МӮЖYҗхҗF‘МҒiӮаӮөӮӯӮНZҗхҗF‘МӮЖWҗхҗF‘МҒjӮЖӮМҠФӮЙӮНӢ@”\“IҒi‘¶ҚЭӮөӮДӮўӮйҲв“`ҺqҒjӮЙ‘еҚ·ӮНӮИӮўӮЖҚlӮҰӮзӮкҒAҗ«•Әү»ӮМҲшӮ«ӢаӮрҲшӮӯҲв“`ҺqӮМӮЭӮӘҲЩӮИӮйӮЖҚlӮҰӮзӮкӮйҒB
ҒңӮИӮәғҒғ_ғJӮЕҗ«ҢҲ’иҲв“`ҺqӮӘ“Ҝ’иӮЕӮ«ӮҪӮ©
Ғ@Ғ@ҡM“ы—ЮҲИҠOӮМҗТ’Е“®•ЁӮЙӮЁӮҜӮйҗ«ҢҲ’иҲв“`Һq“Ҝ’иӮМҺҺӮЭӮНҒAӮЩӮЖӮсӮЗӮӘҺё”sӮЙҸIӮнӮБӮҪҒBӮұӮМӮжӮӨӮИҸуӢөүәҒAүдҒXӮӘғҒғ_ғJҗ«ҢҲ’иҲв“`Һq“Ҝ’иӮЙҗ¬ҢчӮөӮҪ—қ—RӮНҒAғҒғ_ғJӮӘ–{—ҲӮаӮБӮДӮўӮҪҺАҢұ“®•ЁӮЖӮөӮДӮМ“Бҗ«ӮрҠҲӮ©ӮөӮҪӮұӮЖӮЙӮ ӮйҒB
1.Ғ@җ«ӮӘҲв“`“IӮЙҢҲӮЬӮйӮұӮЖ
Ғ@Ӣӣ—ЮӮМҗ«ҢҲ’и—lҺ®ӮНӮҙӮЬӮҙӮЬӮЕӮ ӮйҒBғҒғ_ғJӮМӮжӮӨӮЙҗ«ӮӘҲв“`“IӮЙҢҲ’иӮіӮкӮДӮўӮйҺнӮ©ӮзҒAҠВӢ«—vҲцӮЙӮжӮиҗ«ӮӘҢҲ’иӮіӮкӮДӮўӮйҺнӮЬӮЕӮ ӮйҒB”ӯҗ¶җ¶•ЁҠwӮМғӮғfғӢҗ¶•ЁӮЖӮөӮД‘Ҫ—pӮіӮкӮДӮўӮйғ[ғuғүғtғBғbғVғ…ӮЕӮНҒAғQғmғҖӮМ‘Sү–Ҡо”z—сӮӘ–ҫӮзӮ©ӮЖӮИӮиӮВӮВӮ ӮйӮӘҒAҗ«ҢҲ’и—lҺ®ӮіӮҰ•s–ҫӮЕӮ ӮйҒB
Ғ@ғҒғ_ғJӮМҗ«ӮӘҲв“`“IӮЙҢҲӮЯӮзӮкӮДӮўӮйӮұӮЖӮНҒAҳр“c—ҙ—Y (1921) ӮӘ‘МҗFӮМү©җFҒi”кҗFҒjӮрҢҲӮЯӮйҲв“`ҺqӮМҢАҗ«Ҳв“`Ӯр”ӯҢ©ӮөӮҪӮұӮЖӮЙӮжӮи–ҫӮзӮ©ӮЖӮИӮБӮҪҒBd-rRҢn“қӮЕӮНrҲв“`ҺqҚАӮМ—тҗ«‘О—§Ҳв“`ҺqӮ’ӮӘXҗхҗF‘МҸгӮЙҒA—Dҗ«‘О—§Ҳв“`ҺqRӮӘYҗхҗF‘МҸгӮЙ‘¶ҚЭӮ·ӮйҢӢүКҒAXXҢВ‘МҒiҲв“`ҺqҢ^rrҒjӮН‘МҗFӮӘ”’җFӮЕҺ“ҒAXYҢВ‘МҒiҲв“`ҺqҢ^ӮqrҒjӮН‘МҗFӮӘү©җFӮЕ—YӮЖӮИӮйҒBӮөӮҪӮӘӮБӮДҒAӮұӮМҢn“қӮЕӮНҒAҲв“`“IӮИҗ«Ӯр‘МҗFӮЕ”»’иӮЕӮ«ӮйҒBӮұӮМҢn“қӮр—pӮўӮҪҗ«“]Ҡ·ӮМҢӨӢҶӮН1950-60”N‘гӮЙҺR–{Һһ’jӮЙӮжӮи‘Ҫҗ”•сҚҗӮіӮкҒAҗ«•Әү»ҢӨӢҶӮМҠо”ХӮрӮВӮӯӮБӮҪҒB
2.Ғ@ӢЯҢрҢnӮМ‘¶ҚЭ
Ғ@ӢЯҢрҢnӮНҒAҢZ–…Ңр”zӮр20җў‘гҲИҸгҢJӮи•ФӮ·ӮұӮЖӮЙӮжӮи“ҫӮзӮкӮйҲв“`“IӮЙӢПҲкӮИҢn“қӮЕӮ ӮйҒB•ъҺЛҗьҠҙҺуҗ«ӮИӮЗҢВ‘МҠФӮМӮОӮзӮВӮ«ӮМ‘еӮ«ӮўҲв“`Ң`ҺҝӮӘӢПҲкӮЙӮИӮйӮұӮЖӮЙүБӮҰӮДҒAғQғmғҖӮМү–Ҡо”z—сӮр’ІӮЧӮҪҺһӮЙ‘ҪҢ^ӮрҚl—¶ӮөӮИӮӯӮДӮжӮўӮұӮЖӮаҲв“`Ҡw“IҢӨӢҶӮЙӮН—LҢшӮЕӮ ӮйҒBғQғmғҖғVҒ[ғNғGғ“ғXҢvүжӮаӢЯҢрҢnӮМҲкӮВӮЕӮ ӮйHd-rRҢn“қӮр—pӮўӮДӮИӮіӮкӮҪ2)ҒBӮұӮМӮжӮӨӮИӢЯҢрҢnӮӘ—ҳ—pӮЕӮ«ӮйҗТ’Е“®•ЁӮНҡM“ы—ЮҲИҠOӮЕӮНғҒғ_ғJӮҫӮҜӮЕӮ ӮйҒB
3.Ғ@‘еӮ«ӮИҺн“а•ПҲЩ
Ғ@“ъ–{ҺYӮМ–мҗ¶ғҒғ_ғJӮНҒA–k“ъ–{ҸW’cҒA“м“ъ–{ҸW’cӮЖӮўӮӨӮQӮВӮМ’nҲжҸW’cӮЙ•ӘӮҜӮзӮкӮйҒBӮұӮМ—јҸW’cӮНҲв“`“IӮЙ‘еӮ«ӮӯҲЩӮИӮБӮДӮўӮйҒBғ~ғgғRғ“ғhғҠғAғQғmғҖӮМү–Ҡо”z—сӮЕ8.9Ғ“3)ҒAҠjғQғmғҖӮЕ3.4Ғ“2)ӮМҲбӮўӮӘӮ ӮйҒBӮіӮзӮЙҒAӮ»ӮкӮјӮкӮМҸW’cӮ©ӮзӢЯҢрҢnӮӘҺч—§ӮөӮДӮЁӮиҒAӮұӮкӮзӮМӢЯҢрҢnӮМҠФӮЕӮНҺ©—RӮЙҢр”zӮӘӮЕӮ«ӮйҒBҸЪҚЧӮИҲв“`’nҗ}ӮрҚ\’zӮ·ӮйӮҪӮЯӮЙ•Kҗ{ӮМDNAғ}Ғ[ғJҒ[Ӯр“ҫӮйӮЙӮНҒAҢn“қҠФӮМ‘ҪҢ^ӮӘ•K—vӮЕӮ ӮйҒBғҒғ_ғJӮМҸкҚҮҒA–k“ъ–{ҸW’c—R—ҲӮМӢЯҢрҢnӮЖ“м“ъ–{ҸW’c—R—ҲӮМӢЯҢрҢnӮМҠФӮЕӮНҒAғQғmғҖ100ү–ҠоӮр”дҠrӮ·ӮкӮОҒA3ҢВ’ц“xӮМү–Ҡо’uҠ·Ғi‘ҪҢ^ҒjӮӘҢҹҸoӮЕӮ«ӮйӮЖӮўӮӨҢvҺZӮЙӮИӮиҒA—eҲХӮЙDNAғ}Ғ[ғJҒ[ӮӘ“ҫӮзӮкӮйҒB
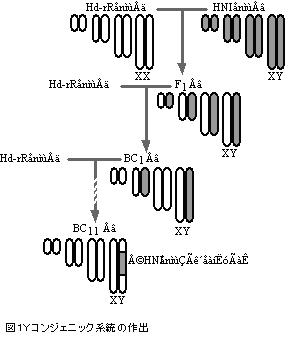
4.Ғ@ӮxғRғ“ғWғFғjғbғNғҒғ_ғJ
Ғ@ӮұӮкӮЬӮЕӮЙҸqӮЧӮҪғҒғ_ғJӮМ“Б’ҘӮрҗ«ҢҲ’иҲв“`Һq“Ҝ’иӮМӮҪӮЯӮЙҚЕ‘еҢАӮЙҠҲ—pӮ·ӮйӮМӮрүВ”\ӮЙӮөӮҪӮМӮӘҒAYғRғ“ғWғFғjғbғNғҒғ_ғJҢn“қӮМҚмҸoӮЕӮ ӮБӮҪҒBYғRғ“ғWғFғjғbғNҢn“қӮНҒA“м“ъ–{ҸW’c—R—ҲӮМӢЯҢрҢnHd-rRӮМҺ“ӮЖ–k“ъ–{ҸW’c—R—ҲӮМӢЯҢрҢnHNIӮМ—YӮЖӮрҢр”zӮөӮД“ҫӮҪF1—YӮрHd-rRҺ“ӮЦ–ЯӮөҢр”zӮ·ӮйӮұӮЖӮрҢJӮи•ФӮ·ӮұӮЖӮЙӮжӮиҒAHd-rRӮМҲв“`“I”wҢiӮЙҒAHNIӮМҗ«ҢҲ’и—МҲжҒiYҗхҗF‘МӮМҲк•”ҒjӮр“ұ“ьӮөӮҪӮаӮМӮЕӮ ӮйҒiҗ}1ҒjҒBӮұӮМҢn“қӮНҒAd-rRҢn“қӮЖ“Ҝ—lӮЙҲв“`“IӮИҗ«Ӯр‘МҗFӮ©Ӯз”»’иӮЕӮ«ӮйҒBӮЬӮҪҒAHNI—R—ҲӮМғQғmғҖӮНYҗхҗF‘МҸгӮМҗ«ҢҲ’и—МҲжҺь•УӮМӮЭӮЙ‘¶ҚЭӮ·ӮйҒBӮВӮЬӮиҒAӮұӮМҢn“қӮМ—YӮМғQғmғҖӮЕӮНҒAҗ«ҢҲ’и—МҲжҺь•УӮМӮЭӮӘHd-rRӮЖHNIӮМғwғeғҚҗЪҚҮӮМҸу‘ФӮЙӮИӮБӮДӮўӮйҒBӮөӮҪӮӘӮБӮДҒADNAғ}Ғ[ғJҒ[ӮМҲв“`ҺqҢ^ӮӘғwғeғҚӮЕӮ ӮкӮОҒAӮұӮМғ}Ғ[ғJҒ[ӮНҗ«ҳAҚҪӮЕӮ ӮйӮЖҢҫӮҰӮй4)ҒB
ҒңғҒғ_ғJҗ«ҢҲ’иҲв“`ҺqӮМ“Ҝ’и
Ғ@Ғ@Ғ@Ӯ·ӮЕӮЙғҒғ_ғJҗ«ҢҲ’иҲв“`Һq“Ҝ’иӮЙҠЦӮ·Ӯй‘ҚҗаӮӘ5,6)Ӯ ӮйӮМӮЕҒAӮұӮұӮЕӮНҠИ’PӮЙҸqӮЧӮйҒB
1.Ғ@ғ|ғWғVғҮғiғӢғNғҚҒ[ғjғ“ғOӮМӮҪӮЯӮМҸҖ”хҒ@(Ӯ»ӮМӮP)
Ғ@ғҒғ_ғJӮМҗ«ҢҲ’иҲв“`ҺqӮНҗ«җхҗF‘МҸгӮЙ‘¶ҚЭӮ·ӮйӮӘҒAӢп‘М“IӮИӢ@”\Ӯв‘Ҡ“ҜҲв“`ҺqӮМ‘¶”ЫӮН•s–ҫӮЕӮ ӮБӮҪҒBӮұӮМӮжӮӨӮЙҗі‘М•s–ҫӮИҲв“`ҺqӮр“Ҝ’иӮ·ӮйӮЙӮНҒAҗхҗF‘МҸгӮМҲК’uҸо•сӮрӮаӮЖӮЙҲв“`ҺqӮр“Ҝ’иӮ·ӮйҒuғ|ғWғVғҮғiғӢғNғҚҒ[ғjғ“ғO–@ҒvӮӘҚЕ“KӮЕӮ ӮйҒB
Ғ@ӮұӮкӮНҒA•\Ң»Ң^Ӯ©ӮзҲв“`ҺqӮрғNғҚҒ[ғjғ“ғOӮ·Ӯй•ы–@ӮЕҸҮҲв“`ҠwӮМүӨ“№ӮЕӮ ӮйӮӘҒA“–ҺһғҒғ_ғJӮЕӮНӮЬӮҫҗ¬Ңч—бӮӘӮИӮ©ӮБӮҪҒBӮ»ӮкӮН‘еӮ«ӮИғCғ“ғTҒ[ғgӮрҠЬӮЮғQғmғҖғүғCғuғүғҠҒ[ӮӘ‘¶ҚЭӮөӮИӮ©ӮБӮҪӮ©ӮзӮЕӮ ӮйҒBӮ»ӮұӮЕӮЬӮёҒA‘еӮ«ӮИғCғ“ғTҒ[ғgӮр•ЫҺқӮЕӮ«ӮйҚЧӢЫҗlҚHҗхҗF‘МҒiBACҒjғxғNғ^Ғ[Ӯр—pӮўӮҪғQғmғҖғүғCғuғүғҠҒ[ӮрҢcүһ‘еҠwӮМҗуҗмҸCҲкҸ•ҺиҒAҗҙҗ…җMӢ`ӢіҺцҒA–јҢГү®‘еҠwӮМ–xҠ°ӢіҺцӮзӮМӢҰ—НӮр“ҫӮДҚмҗ¬ӮөӮҪ1, 7)ҒBBACғүғCғuғүғҠҒ[ӮЖҗ«ҳAҚҪDNAғ}Ғ[ғJҒ[ӮӘӮ ӮкӮОҒAҗхҗF‘М•аҚsӮрҢJӮи•ФӮ·ӮұӮЖӮЕҒAҲв“`’nҗ}Ӯр•Ё—қ’nҗ}ӮЖ‘ОүһӮГӮҜӮДӮўӮҜӮйҒB
Ғ@BACғүғCғuғүғҠҒ[Қ\’zӮЙүдҒXӮӘ—pӮўӮҪғQғmғҖDNAӮМғ\Ғ[ғXӮЙҒgғ~ғ\ҒhӮӘӮ ӮйҒBӮ»ӮкӮНҒAҸгҸqӮМYғRғ“ғWғFғjғbғNҢn“қӮМ—YӮр—pӮўӮҪӮұӮЖӮЕӮ ӮйҒBӮұӮМҢn“қӮМ—YӮНҒAXҗхҗF‘МӮӘHd-rR—R—ҲҒA YҗхҗF‘МӮӘHNI—R—ҲӮМӮҪӮЯҒAXҗхҗF‘МӮЖYҗхҗF‘МӮрӢж•КӮөӮИӮӘӮзҗхҗF‘М•аҚsҒiҢгҸqҒjӮрӮ·ӮйӮұӮЖӮӘӮЕӮ«ӮйӮ©ӮзӮЕӮ ӮйҒB
2.Ғ@ғ|ғWғVғҮғiғӢғNғҚҒ[ғjғ“ғOӮМӮҪӮЯӮМҸҖ”хҒ@(Ӯ»ӮМӮQ)
Ғ@ӮаӮӨҲкӮВҒAҗ«ҢҲ’иҲв“`ҺqӮМғ|ғWғVғҮғiғӢғNғҚҒ[ғjғ“ғOӮЙ•K—vӮИ“№ӢпӮНҒAҗ«ҢҲ’и—МҲжҺь•УӮЕӮМ‘gҠ·ҢВ‘МӮЕӮ ӮйҒBҗ«ҢҲ’иҲв“`ҺqӢЯ–TӮЕӮМ‘gҠ·ҢВ‘МӮӘ‘ҪӮҜӮкӮО‘ҪӮўӮЩӮЗҒAҗ«ҢҲ’иҲв“`ҺqӮрҸЪҚЧӮЙғ}ғbғvӮЕӮ«ӮйҒBӮөӮ©ӮөҒAXY—YӮЕӮНYҗхҗF‘МӮМҗ«ҢҲ’и—МҲжӮрҠЬӮЮ‘еӮ«ӮИ—МҲжӮЕXҗхҗF‘МӮЖӮМҠФӮЕ‘gҠ·ӮӘӢNӮұӮзӮИӮ©ӮБӮҪҒBҲк•ыҒAҗ«“]Ҡ·ӮөӮҪXYҺ“ӮЕӮНҒAӮ Ӯй’ц“x‘gҠ·ӮӘӢNӮұӮйӮұӮЖӮӘ•ӘӮ©ӮБӮДӮўӮҪ8)ҒBӮ»ӮұӮЕҒAӮxғRғ“ғWғFғjғbғNҢn“қӮ©Ӯзҗ«“]Ҡ·XYҺ“Ӯр“ҫҒAӮ»ӮМҺq‘·җ”–ң”цӮ©Ӯз‘gҠ·ҢВ‘МӮр’TҚхӮөӮҪҒB
3.Ғ@җхҗF‘М•аҚs
Ғ@ҢuҢхin situғnғCғuғҠғ_ғCғ[Ғ[ғVғҮғ“–@ӮЙӮжӮйҗ«җхҗF‘МӮМ“Ҝ’и4)ӮЖҒA“®Ңҙ‘М’nҗ}9)Ӯ©ӮзҒAғҒғ_ғJӮМҗ«җхҗF‘МӮНҗхҗF‘МӮМ’ҶӮЕҲк”Ф‘еӮ«ӮўҺҹ’Ҷ•”“®Ңҙ‘МҢ^җхҗF‘МӮМҲкӮВӮЕҒAҗ«ҢҲ’и—МҲжӮНҒAӮ»ӮМ’·ҳrӮЙ‘¶ҚЭӮ·ӮйӮұӮЖӮӘ•ӘӮ©ӮБӮДӮўӮҪҒBҗ«ҢҲ’и—МҲжӮЖ–с1ғZғ“ғ`ғӮғӢғKғ“ҒicMҒA100ҢВ‘МӮЙӮPҢВ‘МӮМ‘gҠ·ҢВ‘МӮӘ“ҫӮзӮкӮйҒjӮМҲв“`Ӣ——ЈӮЙғ}ғbғvӮіӮкӮДӮўӮҪDNAғ}Ғ[ғJҒ[ӮЕӮ ӮйSL110)ӮН’·ҳrӮМҗ«ҢҲ’и—МҲжӮжӮи“®Ңҙ‘М‘ӨӮЙҲК’uӮөӮДӮўӮҪҒBӮЬӮҪҒAғeғҚғҒғA‘ӨӮЙӮН‘МҗFӮМӮ’Ҳв“`ҺqҚАӮӘғ}ғbғvӮіӮкӮДӮўӮҪҒBӮ»ӮұӮЕҒASL1ӮжӮиҗхҗF‘М•аҚsӮрҠJҺnӮөӮҪҒBҺиҸҮӮМҠT—ӘӮНҲИүәӮМӮжӮӨӮЕӮ ӮйҒiҗ}2ҒjҒB
Ғ@ӮЬӮёҒASL1ӮрғvғҚҒ[ғuӮЙBACғүғCғuғүғҠҒ[ӮрғXғNғҠҒ[ғjғ“ғOӮөҒASL1ӮрҠЬӮЮBACғNғҚҒ[ғ“Ӯр“ҫӮйҒBӮұӮМBACғNғҚҒ[ғ“ӮМ––’[”z—сӮжӮиҗVӮҪӮИDNAғ}Ғ[ғJҒ[ӮрҚмӮйӮұӮЖӮӘӮЕӮ«ӮйҒBӮұӮМDNAғ}Ғ[ғJҒ[ӮрғvғҚҒ[ғuӮЖӮөӮДҚДӮСBACғүғCғuғүғҠҒ[ӮрғXғNғҠҒ[ғjғ“ғOӮ·ӮйӮЖӮЖӮаӮЙҒA‘gҠ·ҢВ‘МӮМҲв“`ҺqҢ^Ӯр’ІӮЧӮйӮұӮЖӮЕҒAҗ«ҢҲ’и—МҲжӮМҸЪҚЧӮИҲв“`’nҗ}ӮрҚмҗ¬ӮЕӮ«ӮйҒBSL1Ӯ©ӮзӮWүс–ЪӮМҗхҗF‘М•аҚsӮМҢӢүК“ҫӮҪDNAғ}Ғ[ғJҒ[ 51H7.F ӮӘҒAҗ«ҢҲ’иҲв“`ҺqӮрӢІӮсӮЕSL1ӮМ”Ҫ‘О‘ӨӮЙ’BӮөӮҪӮұӮЖӮЕҒAҗ«ҢҲ’иҲв“`ҺqӮрҠЬӮЮ—МҲжӮрғJғoҒ[Ӯ·ӮйBACғRғ“ғeғBғOӮӘҠ®җ¬ӮөӮҪҒBӮұӮМҺһ“_ӮЕҒAғҒғ_ғJYҗхҗF‘МӮМҗ«ҢҲ’и—МҲжӮНӮSӮВӮМBACғNғҚҒ[ғ“ӮЕғJғoҒ[ӮіӮкӮҪҒBӮВӮЬӮиҒAӮұӮМӮSӮВӮМBACғNғҚҒ[ғ“ӮМ’ҶӮЙҗ«ҢҲ’иҲв“`ҺqӮН‘¶ҚЭӮ·ӮйӮНӮёӮЕӮ ӮйҒB
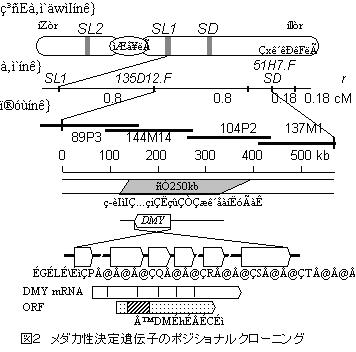
4.Ғ@YҗхҗF‘МӮМҗ«ҢҲ’и—МҲжӮрҢҮҺёӮөӮҪXYҺ“ҢВ‘МӮМ”ӯҢ©
Ғ@ҺҹӮМҚмӢЖӮНҒAӮұӮМ—МҲжӮ©Ӯзҗ«ҢҲ’иҲв“`ҺqӮМҢу•вҲв“`ҺqӮр’TӮ·ӮұӮЖӮЕӮ ӮйҒBcDNAғZғҢғNғVғҮғ“ҒAғGғNғ\ғ“ғgғүғbғvӮИӮЗӮМ•ы–@ӮаҚlӮҰӮзӮкӮҪӮӘҒAҚЕӮаҢӢүКӮӘ‘ҒӮӯҸoӮйӮЖ—\‘zӮөӮҪҒuҗ«ҢҲ’и—МҲжӮМ‘Sү–Ҡо”z—сҢҲ’иҒvӮрҚsӮӨӮұӮЖӮЙӮөӮҪҒBӮSӮВӮМBACғNғҚҒ[ғ“ӮМү–Ҡо”z—сӮрғVғҮғbғgғKғ“ғVҒ[ғNғGғ“ғX–@ӮЙӮжӮиҢҲ’иӮөӮҪҢӢүКҒAӮұӮМ—МҲжӮӘ–с530kbӮЕӮ ӮйӮұӮЖӮӘ•ӘӮ©ӮБӮҪҒB‘Sү–Ҡо”z—сҢҲ’и’ј‘OҒiғhғүғtғgғVҒ[ғNғGғ“ғXӮН“ҫӮДӮўӮҪҺһҒjӮЙҒAҺ©‘R”ӯҗ¶ӮМҗ«“]Ҡ·XYҺ“ӮЖҺvӮнӮкӮйғҒғ_ғJӮрYғRғ“ғWғFғjғbғNҢn“қӮМ’ҶӮЙ”ӯҢ©ӮөӮҪҒBӮұӮМҢВ‘МӮМYҗхҗF‘МӮрҸЪҚЧӮЙ’ІӮЧӮҪӮЖӮұӮлҒAӮSӮВӮМBACғNғҚҒ[ғ“ӮЙғJғoҒ[ӮіӮкӮҪҗ«ҢҲ’и—МҲжӮМӮӨӮҝ”ј•ӘӮЩӮЗӮМ—МҲжҒi–с250kbҒjӮӘҢҮҺёӮөӮДӮўӮйӮұӮЖӮӘ”»–ҫӮөӮҪҒBӮВӮЬӮиҒAӮұӮМXYҢВ‘МӮНҒA250kbӮМ—МҲжӮрҢҮӮўӮҪӮҪӮЯӮЙ—YӮЙ•Әү»ӮЕӮ«ӮёӮЙҺ“ӮЙӮИӮБӮҪӮЖҚlӮҰӮзӮкӮҪҒBӮжӮБӮДҒAӮұӮМ250kbӮМ—МҲжӮЙҗ«ҢҲ’и—МҲжӮНҚiӮиҚһӮЬӮкӮҪҒBҢгӮЙ–ҫӮзӮ©ӮЖӮИӮБӮҪӮұӮЖӮҫӮӘҒAӮұӮМҢҮ‘№ӮөӮҪ—МҲжӮұӮ»ӮӘҒAYҗхҗF‘М“БҲЩ“IӮИ—МҲжӮЕӮ ӮБӮҪҒB
5.Ғ@Ҳв“`ҺqӮМ—\‘Ә
Ғ@ғVғҮғbғgғKғ“ғVҒ[ғNғGғ“ғXӮ©Ӯз“ҫӮзӮкӮҪү–Ҡо”z—сғfҒ[ғ^ӮрӮаӮЖӮЙҲв“`Һq—\‘ӘғvғҚғOғүғҖӮЕ’TҚхӮөӮҪӮЖӮұӮлҒAӮұӮМ—МҲжӮЙ27Ҳв“`ҺqӮӘ—\‘ӘӮіӮкӮҪӮМӮЕҒAӮ»ӮМӮ·ӮЧӮДӮЙӮВӮўӮДҒART-PCR–@ӮЙӮжӮи”ӯҢ»Ӯр’ІӮЧӮҪҢӢүКҒAӮPӮВӮМҲв“`ҺqӮӘҒAҗ«•Әү»ҺһӮЕӮ Ӯйӣzү»ҺһӮЙ—Y“БҲЩ“IӮЙ”ӯҢ»ӮөӮДӮўӮҪҒBRACE–@ӮЙӮжӮиӮұӮМҲв“`ҺqӮМ‘S’·ӮрғNғҚҒ[ғjғ“ғOӮөҒAғAғ~ғmҺ_”z—сӮрҗ„’иӮөӮҪӮЖӮұӮлҒAҒuDMғhғҒғCғ“ҒvӮЖҢДӮОӮкӮй“Б’Ҙ“IӮИғAғ~ғmҺ_”z—с—ЮҺ—ӮМ—МҲжӮӘҠЬӮЬӮкӮДӮўӮҪҒBDMғhғҒғCғ“ӮНғVғҮғEғWғҮғEғoғGӮМҗ«•Әү»ӮЙҠЦӮнӮйdouble sex (dsx) Ҳв“`ҺqӮЖҗь’ҺӮМҗ«•Әү»ӮЙҠЦӮнӮйmale abnormal-3 (mab-3) Ҳв“`ҺqӮЙӢӨ’КӮЙҢ©ӮўӮҫӮіӮкӮҪғAғ~ғmҺ_”z—сӮЕӮ ӮйҒBӮ»ӮұӮЕҒAӮұӮМҲв“`ҺqӮЙҒuYҗхҗF‘МӮМDMғhғҒғCғ“Ҳв“`ҺqҒvӮЖӮўӮӨӮұӮЖӮЕDMYӮЖ–ј‘OӮрӮВӮҜӮҪ1)ҒBҒiӮЩӮЪ“ҜҺһҠъӮЙғҒғ_ғJӮЕDMғhғҒғCғ“Ҳв“`ҺqӮМ‘Ҡ“ҜҲв“`ҺqӮр’TҚхӮөӮДӮўӮҪғhғCғcӮМManfred Schartl”ҺҺmӮМғOғӢҒ[ғvӮаӮұӮМҲв“`ҺqӮЙӮҪӮЗӮи’…ӮўӮДdmrt1bYӮЖ–ј‘OӮр•tӮҜӮДӮўӮйҒBҒj
6.Ғ@җ«ҢҲ’иҲв“`ҺqӮЕӮ ӮйӮұӮЖӮМҸШ–ҫ
Ғ@җ«ҢҲ’и—МҲжӮ©Ӯз”ӯҢ©ӮіӮкӮҪDMYӮНғҒғ_ғJҗ«ҢҲ’иҲв“`ҺqӮМҚЕ—L—НҢу•вӮЕӮ ӮйҒBӮұӮМҲв“`ҺqӮӘҗ«ҢҲ’иҲв“`ҺqӮЕӮ ӮйӮЖӮўӮӨӮұӮЖӮрҺҰӮ·ӮҪӮЯӮЙӮНҒALoss-of- functionӮМҺАҢұӮЖGain-of-functionӮМҺАҢұӮӘ•K—vӮЕӮ ӮйҒB‘OҺТӮН DMYӮН‘¶ҚЭӮ·ӮйӮМӮЙҒA Ӯ»ӮМӢ@”\ҢҮ‘№ӮЕ—YӮЙӮИӮзӮИӮўӮұӮЖҒAҢгҺТӮНDMYӮМӮИӮўҲв“`“IҺ“ (XX) ҢВ‘МӮЙDMYӮр“ұ“ьӮөӮД—YӮЙӮИӮйӮұӮЖӮрҺҰӮ№ӮОӮжӮўӮнӮҜӮЕӮ ӮйҒB
Ғ@ғҒғ_ғJӮМҸкҚҮҒAғGғXғgғҚғQғ“ҸҲ—қӮЙӮжӮи—U“ұӮіӮкӮҪXYҺ“ӮНҒAҗіҸнҺ“ӮЖӮИӮсӮз•ПӮнӮиӮИӮӯҗ¬ҸnӮөҒAҺуҗёүВ”\ӮИ—‘ӮрҺYӮЮӮұӮЖӮ©ӮзҒADMYҲв“`ҺqӮӘӢ@”\ӮөӮИӮўҸкҚҮҒAӮ»ӮМҢВ‘МӮНҺ“ӮЙ•Әү»Ӯ·ӮйӮҫӮҜӮЕӮ»ӮМҢВ‘МӮМҗ¶ҺҖӮвҗ¶җB”\—НӮЙӮНүeӢҝӮөӮИӮўӮЖҚlӮҰӮзӮкӮйҒBӮ»ӮкӮИӮзӮОҒA–мҗ¶ҸW’cӮЙӮЁӮўӮДғүғ“ғ_ғҖӮЙҗ¶Ӯ¶ӮҪ“Л‘R•ПҲЩ’ҶӮЙҒAҗ«ҢҲ’иҲв“`ҺqӮМ“Л‘R•ПҲЩӮН”дҠr“IҚӮ•p“xӮЙҺcӮБӮДӮўӮйӮМӮЕӮНӮИӮўӮ©ӮЖ—\‘zӮөҒA–мҗ¶ғҒғ_ғJӮ©ӮзDMYҲв“`ҺqӮМ“Л‘R•ПҲЩ‘МӮр’TҚхӮ·ӮйӮұӮЖӮЙӮөӮҪҒBӢп‘М“IӮЙӮНҒA–мҗ¶ғҒғ_ғJӮМ•\Ң»Ң^ӮМҗ«•КӮЖDMYӮМ—L–іӮЖӮМҠЦҢWӮр’ІӮЧӮҪҒBӮ»ӮМҢӢүКҒAҗVҠғҢ§ӢҢ”’ҚӘҺsӮЖ•ҹҲдҢ§ӢҢҲ°Ңҙ’¬ӮжӮиDMYӮрҺқӮВҺ“Ӯр”ӯҢ©ӮөӮҪҒBҢр”zҺАҢұӮЙӮжӮиҸЪӮөӮӯ’ІӮЧӮДӮЭӮйӮЖҒA”’ҚӘӮМғҒғ_ғJӮЕӮНDMYӮМ”ӯҢ»—КӮӘҢёҸӯӮөӮДӮЁӮиҒAӮұӮМYҗхҗF‘МӮрҺуӮҜҢpӮўӮҫҺq‘·ӮМ–сӮR•ӘӮМӮQӮӘҺ“ӮЙ•Әү»ӮөӮҪҒBҲк•ыҒAҲ°ҢҙӮМғҒғ_ғJӮЕӮН‘ж3ғGғLғ\ғ“ӮЙӮPү–ҠоӮМ‘}“ьӮӘӮ ӮиҒAӮ»ӮМӮҪӮЯӮЙ•sҠ®‘SӮИDMYғ^ғ“ғpғNӮӘӮЕӮ«ӮДӮўӮйӮЖ—\‘zӮіӮкӮҪҒBӮұӮМYҗхҗF‘МӮрҺуӮҜҢpӮўӮҫҢВ‘МӮНҒA‘SӮДҺ“ӮҫӮБӮҪ11)ҒB
Ғ@Gain-of-function ӮМҺАҢұӮЕӮНҒADMYҲв“`ҺqӮрҠЬӮЮBACғNғҚҒ[ғ“Ӯрd-rRҢn“қӮМ1ҚЧ–EҠъӮЙғ}ғCғNғҚғCғ“ғWғFғNғVғҮғ“Ӯ·ӮйӮұӮЖӮЕҒAғgғүғ“ғXғWғFғjғbғNғҒғ_ғJӮрҚмҸoӮөӮҪҒBҗeӮЙӮИӮБӮҪXXҒi”’җFҒjҢВ‘М58ҢВ‘МӮМӮӨӮҝ13ҢВ‘МӮМ“сҺҹҗ«’ҘӮН—YҢ^ӮЕӮ ӮБӮҪҒBӮЬӮҪҒAӮ»ӮМӮӨӮҝӮWҢВ‘МӮ©ӮзӮНҒAҺҹҗў‘гӮр“ҫӮйӮұӮЖӮӘӮЕӮ«ӮҪ12)ҒB
Ғ@ҲИҸгӮ©ӮзҒADMYӮНғҒғ_ғJӮМҗ«ҢҲ’иҲв“`ҺqӮЖӮөӮДӮМ•K—vҸ\•ӘҸрҢҸӮр–һӮҪӮ·ӮұӮЖӮӘ–ҫӮзӮ©ӮЙӮИӮиҒAҒuDMYӮНғҒғ_ғJӮМҗ«ҢҲ’иҲв“`ҺqӮЕӮ ӮйҒvӮЖӮўӮӨҢӢҳ_ӮЙ’BӮ·ӮйӮұӮЖӮӘӮЕӮ«ӮҪҒB
ҒңDMYӮМ‘Ҫ—lҗ«
1.Ғ@DMYӮМӢNҢ№
Ғ@DMYӮӘғҒғ_ғJӮМҗ«ҢҲ’иҲв“`ҺqӮЕӮ ӮйӮЖӮ·ӮйӮЖҒAҺҹӮМӢ»–ЎӮНҒADMYӮНӮўӮВӮЗӮМҢn“қӮЕҗ¶Ӯ¶ӮҪӮМӮ©ӮЖӮўӮӨӢNҢ№ӮМ–в‘иӮЕӮ ӮйҒB
Ғ@ғҒғ_ғJӮЙҚЕӮаӢЯүҸӮИғnғCғiғ“ғҒғ_ғJӮрҺиҺnӮЯӮЙDMYӮр’TӮөӮҪҒBғҒғ_ғJ—pӮМPCRғvғүғCғ}Ғ[ғZғbғgӮЕғnғCғiғ“ғҒғ_ғJӮМғQғmғҖӮ©ӮзDMYӮМ‘қ•қӮЙҗ¬ҢчӮөҒART-PCRӮвRACE–@ӮЙӮжӮиcDNA‘S’·ӮМү–Ҡо”z—сҢҲ’иӮЙӮаҗ¬ҢчӮөӮҪҒBғnғCғiғ“ғҒғ_ғJӮМDMYӮН—YғQғmғҖӮМӮЭӮЙ‘¶ҚЭӮөҒA“–‘RӮ»ӮМ”ӯҢ»Ӯа—YӮЕӮМӮЭҢҹҸoӮіӮкӮҪҒBӮЬӮҪҒAғҒғ_ғJӮЕDMYӮЙҳAҚҪӮөӮҪDNAғ}Ғ[ғJҒ[ӮНҒAғnғCғiғ“ғҒғ_ғJӮЕӮаDMYӮЙҳAҚҪӮөӮДӮўӮҪӮұӮЖӮ©ӮзҒAғnғCғiғ“ғҒғ_ғJӮМYҗхҗF‘МӮНғҒғ_ғJӮМYҗхҗF‘МӮЖ‘Ҡ“ҜӮЕӮ ӮйӮЖҚlӮҰӮзӮкӮҪҒBDMYӮЙ”z—сӮӘҚЕӮаҺ—ӮДӮўӮйҲв“`ҺqӮЕӮ ӮйDMRT1ӮМғAғ~ғmҺ_”z—сӮЖDMYӮМӮ»ӮкӮЖӮр”дӮЧӮДҚмҗ¬ӮөӮҪ•ӘҺqҢn“қҺчӮНҒAғҒғ_ғJӮЖғnғCғiғ“ғҒғ_ғJӮМӢӨ’К‘cҗжӮЕDMYӮӘDMRT1Ӯ©Ӯз•ӘҠтӮөӮҪӮұӮЖӮрҺҰӮөӮҪ13)ҒBӮВӮЬӮиҒADMYӮМӢNҢ№ӮНҗVӮөӮӯҒAғҒғ_ғJӮЙӮІӮӯӢЯүҸӮМҺнӮЙӮМӮЭDMYӮӘ‘¶ҚЭӮ·ӮйӮЖӮўӮӨӮұӮЖӮЕӮ ӮйҒB
2.Ғ@‘јӮМғҒғ_ғJ‘®Ӣӣ—ЮӮМҗ«ҢҲ’иҲв“`Һq
Ғ@Ӯ»ӮМҢгҒAғҒғ_ғJ‘®Ӣӣ—ЮӮМӮРӮЖӮВӮЕӮ ӮйғCғ“ғhғҒғ_ғJҒiO. dancenaҒjӮМҗ«ҢҲ’иӮЙӮВӮўӮДҗVӮөӮў’mҢ©ӮӘ•сҚҗӮіӮкӮДӮўӮй14ҒjҒBDMYӮрӮаӮҪӮИӮўғCғ“ғhғҒғ_ғJӮМҗ«ӮНXX-XYҢ^ӮМҗ«ҢҲ’и—lҺ®ӮЙӮжӮиҢҲӮЬӮйӮаӮМӮМҒAҗ«ҢҲ’иҲв“`ҺqӮМ‘¶ҚЭӮ·ӮйҗхҗF‘МӮНғҒғ_ғJӮМҗ«җхҗF‘М (1”ФҗхҗF‘М) ӮЕӮНӮИӮӯҸнҗхҗF‘М (10”ФҗхҗF‘М) ӮЖ‘Ҡ“ҜӮЕӮ ӮйӮұӮЖӮӘ”»–ҫӮөӮҪҒBӮВӮЬӮиҒAғҒғ_ғJӮЖӮН‘SӮӯ•КӮМҗ«ҢҲ’иҲв“`ҺqӮӘғCғ“ғhғҒғ_ғJӮЕӮНҗ«ҢҲ’иҲв“`ҺqӮЖӮөӮД“ӯӮўӮДӮўӮйӮЖҗ„‘ӘӮіӮкӮйҒB
Ғ@ӮіӮзӮЙҒAғCғ“ғhғҒғ_ғJӮЖӢЯүҸӮЕӮ ӮйғnғuғXғҒғ_ғJҒiO. hubbsiҒjӮМҗ«ҢҲ’и—lҺ®ӮӘZZ-ZWҢ^ӮЕӮ ӮйӮұӮЖӮӘ–ҫӮзӮ©ӮЙӮИӮБӮҪ15ҒjҒBӮөӮ©ӮаҒAӮ»ӮМҗ«җхҗF‘МӮНғҒғ_ғJӮМҸнҗхҗF‘МӮЕӮ Ӯй5”ФҗхҗF‘МӮЖ‘Ҡ“ҜӮЕӮ ӮБӮҪҒBӮұӮМҺнӮЕӮНZҗхҗF‘МӮЖWҗхҗF‘МӮМҠФӮЙӢtҲКӮӘ‘¶ҚЭӮөҒAҗ«җхҗF‘МӮМҢ`‘ФӮа•Әү»ӮөӮДӮўӮҪҒBӮұӮкӮз2ҺнӮМҗ«ҢҲ’иҲв“`ҺqӮӘ–ҫӮзӮ©ӮЙӮИӮкӮОҒAXX-XYҢ^ӮЖZZ-ZWҢ^ҠФӮМҗ«ҢҲ’и—lҺ®ӮМ•ПҚXӮМғҒғJғjғYғҖӮӘ–ҫӮзӮ©ӮЙӮИӮйӮұӮЖӮӘҠъ‘ТӮіӮкӮйҒB
Ғ@–ў”ӯ•\ӮМҺнӮаҠЬӮЯӮДҚlӮҰӮйӮЖҒAғҒғ_ғJӮМӢЯүҸҺнӮЙӮЁӮўӮДӮНҒAҒiғnғCғiғ“ғҒғ_ғJӮрҸңӮўӮДҒjҺн–ҲӮЙ—lҒXӮИҗ«җхҗF‘МӮӘҗiү»ӮөӮДӮўӮйӮжӮӨӮЕӮ ӮйҒBӮЗӮМӮ®ӮзӮў‘Ҫ—lӮИҲв“`ҺqӮӘғҒғ_ғJ—ЮӮМҗ«ҢҲ’иӮЙҠЦӮнӮБӮДӮўӮйӮМӮ©ҒAҚЎҢгӮЗӮсӮИҗVӮөӮўҗ«ҢҲ’иҲв“`ҺqӮӘҢ©ӮВӮ©ӮБӮДӮӯӮйӮМӮ©ҒAҡM“ы—ЮӮМҗ«ҢҲ’иғJғXғPҒ[ғhӮЖӮМҗЪ“_ӮНүҪӮ©ҒAӢ»–ЎӮНҗsӮ«ӮИӮўҒB
DMYӮЖҗ«•Әү»
1.Ғ@ғҒғ_ғJҗ¶җB‘ғӮМҗ«•Әү»
Ғ@ҢВ‘МӮМҗ«ӮрҢҲӮЯӮйҗ¶җB‘ғӮНҗ¶җBҚЧ–EӮЖ‘МҚЧ–EӮЖӮ©ӮзҚ\җ¬ӮіӮкӮйҒBҗ¶җBҚЧ–EӮМӮаӮЖӮЖӮИӮйҺnҢҙҗ¶җBҚЧ–EӮНҒAҗ¶җB‘ғҢ`җ¬ҲжӮ©Ӯз—ЈӮкӮҪӮЖӮұӮлӮЕ•Әү»ӮөҒAҗ¶җB‘ғҢ`җ¬•”ҲКӮЦӮЖҲЪ“®Ӯ·ӮйҒB”ӯҗ¶үЯ’цӮЙӮЁӮўӮДҒAғҒғ_ғJҗ¶җB‘ғӮМҢ`‘Ф“IӮИҺ“—YҚ·ӮНӣzү»‘OӮМҗ¶җBҚЧ–Eҗ”ӮЙҢ»ӮкӮйҒBӮ·ӮИӮнӮҝҒAӣzү»‘OӮМXXҢВ‘МӮЕӮНҗ¶җBҚЧ–EӮМ•Ә—фҠҲҗ«ӮӘҚӮӮЬӮиҗ¶җBҚЧ–Eҗ”ӮӘ‘қүБӮ·ӮйӮМӮЙ‘ОӮөӮДҒAXYҢВ‘МӮЕӮНӮіӮЩӮЗ•Ә—фӮөӮИӮўӮҪӮЯӮЙҒAӣzү»ӮМҺһ“_ӮЕXXҢВ‘МӮНXYҢВ‘МӮМ2Ғ`3”{ӮМҗ¶җBҚЧ–EӮрӮаӮВҒBӮЬӮҪҒAXXҢВ‘МӮЕӮНӣzү»ҢгҒAҢёҗ”•Ә—фӮЙҲЪҚsӮөӮДӮўӮйҗ¶җBҚЧ–EӮрҠПҺ@ӮЕӮ«Ӯй16)ҒB
2.Ғ@DMYӮМ”ӯҢ»
Ғ@in situғnғCғuғҠғ_ғCғ[Ғ[ғVғҮғ“–@ӮЙӮжӮйӮЖҲЪ“®’ҶӮМҺnҢҙҗ¶җBҚЧ–EҺь•УӮМ‘МҚЧ–EҒAҗ¶җB‘ғҢ`җ¬•”ҲКӮМ‘МҚЧ–EӮЕӮНӢӨӮЙDMYӮМ”ӯҢ»ӮНҢҹҸoӮіӮкӮИӮўҒBҲк•ыҒAҗ¶җB‘ғҢ`җ¬•”ҲКӮЙ“һ’BӮөӮҪҗ¶җBҚЧ–EӮрҺжӮиҲНӮЮ‘МҚЧ–EҒiҸ«—ҲӮМғZғӢғgғҠҚЧ–EҒjӮЕӮНҒADMYӮМ”ӯҢ»ӮӘҢҹҸoӮіӮкӮй16)ҒBӮұӮМӮұӮЖӮ©ӮзҒAҗ«ҢҲ’иҲв“`ҺqӮМ”ӯҢ»ӮНҒAҡM“ы—ЮӮМSryӮЖ“Ҝ—lӮЙғZғӢғgғҠҚЧ–EҢn—сӮЕҺnӮЬӮиҒAғZғӢғgғҠҚЧ–E•Әү»ӮЦӮМҲшӮ«ӢаӮрҲшӮўӮДӮўӮйӮЖҚlӮҰӮзӮкӮйҒBӮҪӮҫӮөҒASryӮМ”ӯҢ»ӮӘҲкүЯҗ«ӮЕӮ ӮйӮМӮЙ‘ОӮөӮДҒADMYӮМ”ӯҢ»ӮНҗeӮМҗё‘ғӮЕӮаҢҹҸoӮЕӮ«ӮйҒBҗ«ғzғӢғӮғ“ҸҲ—қӮЙӮжӮиҗ«“]Ҡ·ӮөӮҪDMYӮрҺқӮҪӮИӮўXX—YӮӘҗіҸнӮИҗё‘ғӮрҺқӮБӮДӮўӮйӮұӮЖӮрҚlӮҰӮйӮЖҒAҗeҗё‘ғӮЕӮМDMYӮМ”ӯҢ»ӮНӢ@”\“IӮЙҲУ–ЎӮӘӮИӮўӮМӮ©Ӯа’mӮкӮИӮўҒB
Ғ@ӮЬӮҪҒADMYҲв“`ҺqҸг—¬ӮМ”ӯҢ»җ§Ңд—МҲжӮЙGFPӮЙӮВӮИӮўӮҫғRғ“ғXғgғүғNғgӮр“ұ“ьӮөӮҪғgғүғ“ғXғWғFғjғbғNғҒғ_ғJӮМGFPғVғOғiғӢӮНҒAҗё‘ғӮМ‘МҚЧ–EӮЕҢҹҸoӮЕӮ«ӮйӮҫӮҜӮЕӮИӮӯҒA—‘‘ғӮМҗ¶җBҚЧ–EҺь•УӮМ‘МҚЧ–EӮЕӮаҢҹҸoӮЕӮ«ӮйҒBӮұӮМӮұӮЖӮ©ӮзҒADMYӮМ”ӯҢ»җ§Ңд—МҲжӮЙӮНҒAҗ¶җBҚЧ–EҺь•УӮМ‘МҚЧ–EӮЕ”ӯҢ»Ӯ·ӮйӮЖӮўӮӨғ|ғeғ“ғVғғғӢӮӘӮ ӮйӮЖүрҺЯӮөӮДӮўӮйҒBXXҢВ‘МӮМҗ¶җB‘ғӮЖXYҢВ‘МӮМҗ¶җB‘ғӮЖӮМ—ј•ыӮЙҒADMYӮӘ”ӯҢ»үВ”\ӮИҒuҸкҒvӮН“Ҝ“ҷӮЙ‘¶ҚЭӮөӮДӮўӮйӮӘҒAXYҢВ‘МӮМӮЭӮӘDMYӮрӮаӮВӮҪӮЯҒADMYӮӘ”ӯҢ»ҒEӢ@”\ӮөҒAҗ¶җB‘ғӮрҗё‘ғ•ыҢьӮЦӮЖ“ұӮўӮДӮўӮӯӮМӮҫӮЖҚlӮҰӮДӮўӮйҒiҸј“cӮз–ў”ӯ•\ҒjҒB
3.Ғ@DMYӮМӢ@”\ӮЖҗ¶җBҚЧ–Eҗ”
Ғ@XYӮЕӮ ӮиӮИӮӘӮзӢ@”\ӮөӮИӮўDMYӮрӮаӮВ“Л‘R•ПҲЩ‘МӮНҺ“ӮЙ•Әү»Ӯ·Ӯй17)ҒBҲк•ыҒAғGғXғgғҚғQғ“ҸҲ—қӮЕӮаXYҺ“Ӯр—U“ұӮ·ӮйӮұӮЖӮӘүВ”\ӮЕӮ ӮйҒBӮөӮ©ӮөҒAӮұӮМ—јҺТӮЙӮВӮўӮДҒA’tӢӣҠъӮМҗ¶җBҚЧ–Eҗ”•Пү»Ӯр”дҠrӮ·ӮйӮЖҒAҲбӮўӮӘ”FӮЯӮзӮкӮйҒBDMY“Л‘R•ПҲЩ‘МӮМҗ¶җBҚЧ–Eҗ”ӮНҠо–{“IӮЙҺ“ӮЖ“ҜӮ¶ғpғ^Ғ[ғ“ӮЕ•Пү»ӮөҒAӣzү»‘OӮЙ‘қүБӮөҒAӣzү»ҢгӮНҢёҗ”•Ә—фӮЙҲЪҚsӮ·ӮйҒBҲк•ыҒAғGғXғgғҚғQғ“ҸҲ—қӮөӮҪXYҢВ‘МӮЕӮНҒAҗ¶җBҚЧ–EӮМҗ”ӮНҒAӣzү»ӮөӮҪҺһ“_ӮЕҗіҸнӮИXXҢВ‘МӮЩӮЗ‘ҪӮӯӮИӮӯҒAҗіҸнӮИXYҢВ‘МӮМҗ”ӮЙӢЯӮўҒBӮөӮ©ӮөҒAҗіҸнӮИXXҢВ‘МӮЖ“Ҝ—lӮЙҢёҗ”•Ә—фӮМҺw•WӮЖӮИӮйScp3Ӯр”ӯҢ»Ӯ·Ӯйҗ¶җBҚЧ–EӮӘҠПҺ@ӮЕӮ«Ӯй18)ҒBӣzү»ҢгӮНҗіҸнXYҢВ‘МӮЖ”дҠrӮөӮДҗ¶җBҚЧ–Eҗ”ӮӘҢ°’ҳӮЙ‘қүБӮөҒAXXӮЖ“Ҝ—lӮИҗ¶җB‘ғӮМҢ`‘ФӮЙӮИӮйҒB
Ғ@Ҳк•ыҒADMYӮМғQғmғҖ—МҲжӮр“ұ“ьӮөӮҪXXғҒғ_ғJӮвҒADMYӮрXXгуӮЙӢӯҗ§”ӯҢ»ӮіӮ№ӮҪҸкҚҮӮЙӮНҒAҸүҠъӮ©ӮзXYгуӮЖ“Ҝ—lӮМҗ¶җB‘ғҗ«•Әү»ғpғ^Ғ[ғ“ӮрҺҰӮ·ҢВ‘МӮӘҢ»ӮкӮй12)ҒBӮұӮкӮзӮМҢӢүКӮНҒADMYӮӘҗ¶җBҚЧ–EӮМ•Ә—фҗ§ҢдӮЙ–§җЪӮЙҠЦӮнӮБӮДӮўӮйӮұӮЖӮрҲГҺҰӮөӮДӮўӮйҒB
ҒңғҒғ_ғJӮМҗ«ӮМүВ‘Yҗ«ӮЖҗ«ҢҲ’иҲв“`ҺqӮМҗiү»ҒE‘Ҫ—lҗ«
Ғ@Ғ@–мҗ¶ҸW’cӮ©Ӯз”ӯҢ©ӮөӮҪDMY“Л‘R•ПҲЩ‘МӮМ‘ҪӮӯӮНҗіҸнӮИҒi–мҗ¶Ң^ҒjDMYӮМӮPү–ҠоӮМin/delӮҫӮБӮҪҒBӢ@”\ҢҮ‘№Ң^DMYӮрӮаӮВYҗхҗF‘МӮрүјӮЙY*ӮЖӮ·ӮйӮЖҒAӮұӮМY*җхҗF‘МӮНӢ@”\“IӮЙӮНXҗхҗF‘МӮЖ“ҷүҝӮЕӮ ӮйҒBҢр”zӮЙӮжӮиҚмҸoӮөӮҪY*Y*ӮНҺ“ӮЙҒAY*YӮН—YӮЙ•Әү»Ӯ·Ӯй17)ҒBӮВӮЬӮиҒAӮұӮМ•ПҲЩҢn“қӮЕӮНҒAY*җхҗF‘МӮЖYҗхҗF‘МӮЖӮМҲбӮўӮНӮPү–ҠоӮЕӮ ӮйӮЖӮўӮӨӮұӮЖӮЙӮИӮйҒBҢ»‘¶Ӯ·Ӯй‘ҪӮӯӮМҺнӮМӮӨӮҝӮЙӮНҒAҒuXҗхҗF‘МӮЖYҗхҗF‘МӮЖӮМҲбӮўӮНӮPү–ҠоҒvӮЕӮ ӮйӮЖӮўӮӨӮұӮЖӮӘӮ Ӯи“ҫӮйӮЖҺvӮнӮкӮйҒBӮұӮМӮжӮӨӮИҗхҗF‘МҚ\җ¬ӮМҺнӮЕӮНҒAҗ«ҢҲ’иҲв“`Һq“Ҝ’иӮНӢЙӮЯӮД“пӮөӮӯӮИӮйӮҫӮлӮӨҒBYҗхҗF‘МҸгӮЙXҗхҗF‘МӮЙӮН‘¶ҚЭӮөӮИӮў250kbӮаӮМ‘}“ь”z—сӮМӮ ӮБӮҪғҒғ_ғJӮНҚKү^ӮҫӮБӮҪӮМӮ©ӮаӮөӮкӮИӮўҒBғҒғ_ғJӮМҗ«ӮНҗ«ғzғӢғӮғ““Ҡ—^ӮвҺуҗё—‘ӮМҚӮү·ҸҲ—қ19)ӮИӮЗӮЙӮжӮБӮД—eҲХӮЙ•ПҚXүВ”\ӮЕӮ ӮйҒBӮ»ӮМ—қ—RӮНҒAҺ“—YӮМҲв“`“IӮИҲбӮўӮӘӢЙӮЯӮДҸ¬ӮіӮўӮұӮЖӮЙӮжӮйӮЖӮаҢҫӮҰӮйӮМӮЕӮ ӮйҒB
ҒңҚЎҢгӮМүЫ‘и
Ғ@Ғ@Ғ@ғҒғ_ғJӮМҗ«ҢҲ’иҲв“`ҺqӮӘ–ҫӮзӮ©ӮЖӮИӮБӮҪӮұӮЖӮЕҒAҗТ’Е“®•ЁӮЙӮЁӮҜӮйҗ«ҢҲ’иҲв“`ҺqӮМ‘Ҫ—lҗ«ӮМҺА‘МӮӘ–ҫӮзӮ©ӮЖӮИӮБӮДӮ«ӮҪҒBӮ»ӮМҲк•ыҒAҗё‘ғҢ`җ¬Ӯв—‘‘ғҢ`җ¬ӮИӮЗҗ¶җB‘ғӮМҗ«•Әү»үЫ’цӮНҗТ’Е“®•ЁӮМҠФӮЕ•Ы‘¶җ«ӮӘҚӮӮўӮұӮЖӮ©ӮзҒAҗТ’Е“®•ЁӮЙӢӨ’КӮөӮҪғҒғJғjғYғҖӮӘӮ ӮйӮЖӮаҗ„’иӮЕӮ«ӮйҒBҚЎҢгҒADMYҺ©‘МӮМӢ@”\ӮЖҗё‘ғ•Әү»ӮМҲв“`ҺqғJғXғPҒ[ғhӮрӮВӮИӮ°ӮйӮұӮЖӮӘҸd—vӮИүЫ‘иӮЕӮ ӮйҒB
Ғ@Ҳк•ыҒAғҒғ_ғJӮЙӢЯүҸӮИҺнӮЙҠЦӮөӮДӮНҒAҗ«ҢҲ’иҲв“`Һq“Ҝ’иҢvүжӮӘҗiҚsӮөӮДӮЁӮиҒAӢЯӮўҸ«—ҲҒAӮ»ӮкӮјӮкӮМҺнӮМҗ«ҢҲ’иҲв“`ҺqӮӘҺҹҒXӮЙ–ҫӮзӮ©ӮЖӮИӮйӮұӮЖӮӘҠъ‘ТӮЕӮ«ӮйҒBҗ¶җB‘ғҗ«•Әү»ӮМҲв“`ҺqғJғXғPҒ[ғhӮӘ•Ы‘¶ӮіӮкӮДӮўӮйӮЖӮ·ӮкӮОҒAғJғXғPҒ[ғhӮМ—lҒXӮИ’iҠKӮМҲв“`ҺqӮНҒAӮ»ӮкӮјӮкӮМҺнӮЕҗ«ҢҲ’иҲв“`ҺqӮЖӮөӮД“ӯӮ«“ҫӮйҒBҗVӮҪӮЙ”ӯҢ©ӮіӮкӮйӢЯүҸҺнӮМҗ«ҢҲ’иҲв“`ҺqӮвғҒғ_ғJҺн“аӮЙ‘¶ҚЭӮ·ӮйDMYҲИҠOӮМҗ«•ПҚXҲв“`Һq•ПҲЩ20) ӮИӮЗӮр“Ҝ’иҒEүрҗНӮ·ӮйӮұӮЖӮЙӮжӮБӮДҒAҗТ’Е“®•ЁӮМҗ«•Әү»ӮМҲв“`ҺqғJғXғPҒ[ғhӮӘ•ӮӮ©ӮСҸгӮӘӮБӮДӮӯӮйӮұӮЖӮрҠъ‘ТӮөӮДӮўӮйҒB
ҒңҺУҺ«
Ғ@Ғ@Ғ@–{ҢӨӢҶӮМҗӢҚsӮМӮҪӮЯӮЙӮНҒA‘ҪӮӯӮМ•ыҒXӮМӢҰ—НӮӘ•Kҗ{ӮЕӮөӮҪҒB“БӮЙҠо‘bҗ¶•ЁҠwҢӨӢҶҸҠӮМ’·а_ үГҚFӢіҺцӮЙӮН•ЁҗS—ј–КӮЕғTғ|Ғ[ғgӮөӮД’ёӮ«ӮЬӮөӮҪҒBҠоҗ¶ҢӨӮМ“dҺqҢvҺZӢ@ҺәҒA•ӘҗНҺәӮ©ӮзӮМҺxүҮӮа’ёӮ«ӮЬӮөӮҪҒBӮЬӮҪҒAҗhҢыӮЕ—LҲУӢ`ӮИӢcҳ_ӮрӮөӮДӮўӮҪӮҫӮўӮҪа_Ңы “NӢіҺцӮрҺnӮЯӮЖӮ·ӮйҗVҠғ‘еҠwӮМҢӨӢҶҺәӮМҠF—lӮМҚvҢЈӮНӮҪӮўӮЦӮс‘еӮ«ӮИӮаӮМӮЕӮөӮҪҒBӮұӮМҸкӮрҺШӮиӮДҗSӮжӮиҠҙҺУӮўӮҪӮөӮЬӮ·ҒB
Ғң•¶ҢЈ
1) Matsuda, M., Nagahama, Y., Shinomiya, A., Sato, T., Matsuda, C., Kobayashi, T., Morrey, C. E., Shibata, N., Asakawa, S., Shimizu, N., Hori, H., Hamaguchi, S. and Sakaizumi, M. DMY is a Y-specific DM-domain gene required for male development in the medaka fish. Nature, 417: 559-563 (2002).
2) Kasahara, M., Naruse, K., Sasaki, S., Nakatani, Y.,‘ј. The medaka draft genome and insights into vertebrate genome evolution. Nature, 447: 714-719 (2007).
3) Matsuda, M., Yonekawa, H., Hamaguchi, S. Sakaizumi, M.Ғ@Geographic variation and diversity in the mitochondrial DNA of the medaka, Oryzias latipes, as determined by restriction endonuclease analysis. Zool. Sci., 14: 517-526Ғ@(1997).
4) Matsuda M., Matsuda C., Hamaguchi S., Sakaizumi M.Ғ@Identification of the sex chromosomes of the medaka, Oryzias latipes, by fluorescence in situ hybridization.
Cytogenet. Cell Genet., 82: 257-263 (1998).
5) ҺlӢ{ҲӨҒAа_Ңы“NҒAҺрҗт–һ. ғҒғ_ғJӮМҗ«ҢҲ’иҲв“`ҺqӮЖҗ¶җB‘ғӮМҗ«•Әү»Ғ@җТ’Е“®•ЁӮМҗVӮөӮўҗ«ҢҲ’иҲв“`ҺqDMY. ҚЧ–EҚHҠw 22: 1090-1096 (2003).
6) ’·а_үГҚFҒAҸј“cҸҹ. ғҒғ_ғJӮМҗ«ҢҲ’иҲв“`Һq. ҚЧ–EҚHҠw 25: 364-368 (2006).
7) Matsuda M., Kawato N., Asakawa S., Shimizu N., Nagahama Y., Hamaguchi S., Sakaizumi M., Hori H. Construction of a BAC library derived from the inbred Hd-rR strain of the teleost fish, Oryzias latipes. Genes Genet. Syst., 76: 61-63 (2001).
8) Matsuda M., Sotoyama S., Hamaguchi S., Sakaizumi M.Ғ@Male-specific restriction of recombination frequency in the sex chromosomes of the medaka, Oryzias latipes. Genet. Res. Cambr., 73: 225-231 (1999).
9) Sato T. Yokomizo S., Matsuda M., Hamaguchi S. and Sakaizumi M.Ғ@Gene-Centromere Mapping of Medaka Sex Chromosome using Triploid Hybrids between Oryzias latipes and O. luzonensis. Genetica, 111, No. 1-3: 71-75 (2001)
10) Matsuda, M., Kusama, T., Kurihara, Y., Oshiro, T., Hamaguchi, S. Sakaizumi, M. Isolation of a sex chromosome-specific DNA sequence in the medaka, Oryzias latipes. Genes Genet. Syst., 72: 263-268 (1997).
11) Shinomiya, A, Otake, H., Togashi, K., Hamaguchi, S., Sakaizumi, M. Field survey of sex-reversals in the medaka, Oryzias latipes: genotypic sexing of wild populations. Zool. Sci., 21: 613-619 (2004).
12) Matsuda, M., Shinomiya, A., Kinoshita, M., Suzuki, A., Kobayashi, T., Paul-Prasanth, B., Lau, E-l., Hamaguchi, S., Sakaizumi, M. and Nagahama, Y. DMY induces male development in XX medaka fish. Proc Natl Acad Sci. 104: 3865-3870 (2007).
13) Matsuda, M., Sato, T., Toyazaki, Y., Nagahama, Y., Hamaguchi, S. and Sakaizumi, M. Oryzias curvinotus Has DMY, a Gene That Is Required for Male Development in the Medaka, O. latipes. Zool. Sci., 20: 159-161 (2003).
14) Takehana, Y., Demiyah, D., Naruse, K., Hamaguchi, S. and Sakaizumi, M. Evolution of different Y chromosomes in two medaka species, Oryzias dancena and O. latipes. Genetics 175: 1-6 (2007).
15) Takehana, D., Naruse, K., Hamaguchi, S., Sakaizumi, M. Evolution of ZZ/ZW and XX/XY sex-determination systems in the closely related medaka species, Oryzias hubbsi and O. dancena. Chromosoma, in press.
16) Kobayashi, T., Matsuda, M., Kajiura-Kobayashi, H., Suzuki, A., Saito, N., Nakamoto, M., Shibata, N. and Nagahama, Y. Two DM domain genes, DMY and DMRT1, involved in testicular differentiation and development in the medaka, Oryzias latipes. Dev. Dyn., 231: 518-526 (2004).
17) Otake, H., Shinomiya, A., Matsuda, M., Hamaguchi, S., Sakaizumi, M. Wild-derived XY sex-reversal mutants in the medaka, Oryzias latipes. Genetics, 173: 2083-2090 (2006).
18) Paul-Preasanth, B., Matsuda, M., Lau, E-L., Suzuki, A., Sakai, F., Kobayashi, T. and Nagahama, Y. Knock-down of DMY initiates female pathway in the genetic maleҒ@medaka, Oryzias latipes. Biochem. Biophys. Res. Commun., 351: 815-819 (2006).
19) Sato, T., Endo, T., Yamahira, K., Hamaguchi, S., Sakaizumi, M. Induction of female-to-male sex reversal by high temperature treatment in Medaka, Oryzias latipes. Zool. Sci., 22: 985-988 (2005).
20) Shinomiya, A., Kato M., Yaezawa, M., Sakaizumi, M., Hamaguchi, S. Interspecific hybridization between Oryzias latipes and Oryzias curvinotus causes XY sex reversal. J. Exp. Zool., 305A: 890-896 (2006).
|

